On fallen twigs or moss-covered trunks of
deciduous trees, or on fallen leaves of e. g. Salix and
stems of different herbs, such as Filipendula ulmaria. Also found on fallen
hazel nuts and small woody items in the litter. Early summer to early winter. In milder parts of the region all year. A common and widespread species. Fairly common in southern part of Norway but not recorded in the northernmost counties. Possibly overlooked.
Pileus 2-5 mm
across, hemispherical, conical, becoming more or less parabolical
with age, translucent-striate, shallowly sulcate, white-furfuraceous
to floccose, glabrescent, white or grey. Lamellae
7-13, not always reaching the stipe, ascending, narrowly
adnate or attached to a pseudocollarium, the edge convex,
white. Stipe 5-30 mm long,
filiform, straight to flexuous, equal, greyish-hyaline,
puberulous, glabrescent with age, but often hirsute below,
the base somewhat bulbous with a small, hirsute, white,
basal disc. Odour none. Taste recorded as mild.
Basidia
13-20 x 7-9 µm, clavate or obpyriform, 2- spored.
Spores
(7-)8-9.7 x (4.3)5.5-6(-6.5) µm, Qav 1.4-1.7, broadly pip-shaped to subglobose, smooth, amyloid. Cheilocystidia
9-25 x -12 µm, numerous, apparently forming a sterile band, clavate or obpyriform to fusiform,
mostly with a slender, straight to curved, simple or occasionally
furcate neck up to 20 µm long; smooth or
covered with warts or cylindrical excrescences 0.5-1.5 x
0.5 µm. Pleurocystidia absent. Lamellar trama dextrinoid. Hyphae
of the pileipellis smooth to spinulose,
with terminal
cells globose to obpyriform (acanthocysts), densely covered
with warts and short, cylindrical excrescences, not gelatinous. Caulocystidia
20-110 x 6-10 µm, +/-lanceolate, acute with a widened base, smooth, thin-walled.
Basal disc hyphae
often in chains of 2-4, smooth, terminated by cylindrical, clavate, lageniform cells, obtuse to subacute, smooth
or rarely with a few coarse, apical excrescences. Clamps abundant.
Mycena tenerrima occasionally
grows on hazel nuts. Two other
species have been reported growing on hazel nuts: M.
discopus (Lév.) Quél. and M.
nucicola. M. discopus
is a somewhat dubious species and awaits further description.
Desjardin (1995: 79) excluded it as a nomen dubium.
Huijsman (1958) described the cheilocystidia of M.
nucicola as clavate, lacking an apical
rostrum and Maas Geesteranus (1991b) did not report otherwise.
Desjardin (1995) and Robich (2003), however, showed that
many of the cheilocystidia near the pileus margin have a
single apical projection. This has been confirmed by several Norwegian collections. M. nucicola can be separated from M., tenerrima on account of the
4-spored basidia, narrower spores, absent or very rare clamps, and, most important, spinulose terminal cells of the basal disc hyphae.
Aronsen & Larsson (2015) sequenced collections identified as M. nucicola and M. tenerrima from Norway and Sweden and found them identical, and indicated that M. nucicola should be regarded as a synonym of the latter. This was followed by Aronsen & Læssøe (2016). This apparently was an error. Although there are no distinct macroscopic differences, the two species can be clearly separated on account of microscopic features. The differences have been confirmed by recent ITS sequences.
One problem is connected to M. adscendens var. carpophila (J.E. Lange) Desjardin, which seems
to be very unsufficiently known. The type does not exist. Lange reported the species as having 4-spored
basidia, narrower basidiospores, and fruits on Fagus pericarps, but he did not report on cheilocystidia shape
nor caulocystidia, and he certainly was not aware of microscopic
features of the basal disc. Until new material can be found
it is impossible to tell anything certain about this variety. |
 |
Jagers et al. (2022) found two distinctly different phylogenetic species of M. tenerrima in material from the Netherlands, one probably representing M. nucicola. Deceuninck (2023) published a preliminary study, where she lined up the differences between the two species.
Van den Berg et al. (2000) described the new species Mycena cecidiophila A.P. Berg, Berg-Block, Noordel. & Uljé, that was found growing on old knopper galls on the cups of Quercus robur. It differs from Mycena tenerrima in having a consistently brownish centre of the pileus and a conspicuously fimbriate margin. In addition it was characterized by the absence of pleuro- and cheilocystidia and a negative Melzer-reaction in the lamellar trama. The authors also proposed the new section Cecidiophilae to accomodate the new species. Later, this taxon has been shown to be identical with M. rhenana Maas Geest & Winterh. (Noten & Vannieuwerburgh 2009).
Both M. clavularis and M. corynephora may resemble M. tenerrima. Both have globose spores.
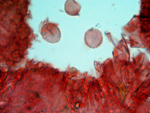
Microphotos of basal disc hyphae and cheilocystidia
|